ALLHORMON.NAROD.RU
Все о гормона - сайт посвященные эндокринной системе человека.
|
Главная |
Добавить в избранное
ГипофизГипофиз регулирует активность ряда желез внутренней секреции и служит местом выделения гипоталамических гормонов крупноклеточных ядер гипоталамуса. Состоит из двух эмбриологически, структурно и функционально различных частей - нейрогипофиза - выроста промежуточного мозга и аденогипофиза, ведущей тканью которого служит эпителий. Аденогидофиз разделяется на более крупную переднюю долю, узкую промежуточную и слабо развитую туберальную часть(рис. 1). 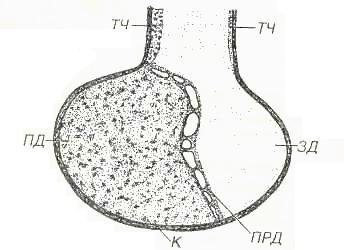 Рис. 1. Гипофиз. ПД - передняя доля, ПРД - промежуточная доля, ЗД - задняя доля, ТЧ - туберальная часть, К - капсула. Гипофиз покрыт капсулой из плотной волокнистой ткани. Его строма представлена очень тонкими прослойками рыхлой соединительной ткани, связанными с сетью ретикулярных волокон, которая в аденогипофизе окружает тяжи эпителиальных клеток и мелкие сосуды. Передняя доля гипофиза у человека составляет около 75% его массы; она образована анастомозирующими тяжами (трабекулами) аденоцитов, тесно связанными с системой синусоидных капилляров. Форма аденоцитов варьирует от овальной до полигональной. На основании особенностей окраски их цитоплазмы выделяют: 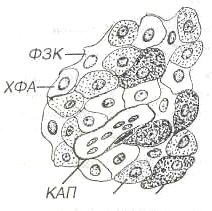 Рис 2. Передняя доля гипофиза. АА - ацидофильные аденоциты, БА - бэзофильные аденоциты, ХФА - хромофобные аденоциты, ФЗК - фолликулярно-звездчатые клетки, КАП - капилляр. 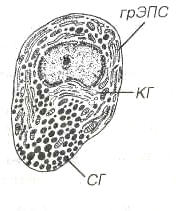 Рис. 3. Ультраструктура соматотропа: грЭПС - гранулярная эндоплазматическэя сеть, КГ - комплекс Гольджи, СГ - секреторные гранулы. 1. Хромофильные аденоциты (хромофилы) характеризуются развитым синтетическим аппаратом и накоплением в цитоплазме секреторных гранул, содержащих гормоны (рис. 3). В зависимости от окраски секреторных гранул хромофилы подразделяют на ацидофилы и базафилы. а) ацидофилы (около 40% всех аденоцитов) - мелкие округлые клетки с хорошо развитыми органеллами и высоким содержанием крупных гранул - включают два типа:
б)базофилы (10-20%) крупнее ацидофилов, однако их гранулы мельче и обычно содержатся в меньшем количестве. Включают гонадотропы, тиротропы и адренокортикотропы:
2. Хромофобные аденоциты (хромофобы) - разнородная группа клеток, которая включает:
Промежуточная доля у человека развита очень слабо и состоит из узких прерывистых тяжей базофильных и хромофобных клеток, которые секретируют МСГ - меланоцитостимулирующий гормон (активирует меланоциты) и ЛПГ - липотропный гормон (стимулирует обмен жиров). МСГ и ЛПГ (как и АКТГ) являются продуктами расщепления ПОМК. Встречаются кистозные полости, выстланные реснитчатыми клетками и содержащие негормональное белковое вещество - коллоид. Туберальная часть в виде тонкого (25-60 мкм) рукава покрывает гипофизарную ножку, отделяясь от нее узким слоем соединительной ткани. Она состоит из тяжей хромофобных и хромофильных клеток; Задняя доля содержит:
Физиология гипофизаГипофизу принадлежит особая роль в системе желез внутренней секреции. С помощью своих гормонов он регулирует деятельность других эндокринных желез. Гормоны передней доли гипофиза.
Первые 4 гормона регулируют функции так называемых периферических желез внутренней секреции. Соматотропин и пролактин сами действуют на ткани-мишени. Адренокортикотропный гормон (АКТГ)Адренокортикотропный гормон (АКТГ), или кортикотропин, оказывает стимулирующее действие на кору надпочечников. В большей степени его влияние выражено на пучковую зону, что приводит к увеличению образования глюкокортикоидов, в меньшей — на клубочковую и сетчатую зоны, поэтому на продукцию минералокортикоидов и половых гормонов он не оказывает значительного воздействия. За счет повышения синтеза белка (цАМФ-зависимая активация) происходит гиперплазия коркового вещества надпочечников. АКТГ усиливает синтез холестерина и скорость образования прегненолона из холестерина. Вненадпочечниковые эффекты АКТГ заключаются в стимуляции липолиза (мобилизует жиры из жировых депо и способствует окислению жиров), увеличении секреции инсулина и соматотропина, накоплении гликогена в клетках мышечной ткани, гипогликемии, что связано с повышенной секрецией инсулина, усилении пигментации за счет действия на пигментные клетки меланофоры. Продукция АКТГ подвержена суточной периодичности, что связано с ритмичностью выделения кортиколиберина. Максимальные концентрации АКТГ отмечаются утром в б — 8 часов, минимальные — с 18 до 23 часов. Образование АКТГ регулируется кортиколиберином гипоталамуса. Секреция АКТГ усиливается при стрессе, а также под влиянием факторов, вызывающих стрессогенные состояния: холод, боль, физические нагрузки, эмоции. Гипогликемия способствует увеличению продукции АКТГ. Торможение продукции АКТГ происходит под влиянием самих глюкокортикоидов по механизму обратной связи. Избыток АКТГ приводит к гиперкортицизму, т.е. увеличенной продукции кортикостероидов, преимущественно глюкокортикоидов. Это заболевание развивается при аденоме гипофиза и носит название болезни Иценко—Кушинга. Основные проявления ее: гипертония, ожирение, имеющее локальный характер (лицо и туловище), гипергликемия, снижение иммунной защиты организма. Недостаток гормона ведет к уменьшению продукции глюкокортикоидов, что проявляется нарушением метаболизма и снижением устойчивости организма к различным влияниям среды. Тиреотропный гормон (ТТГ)Тиреотропный гормон (ТТГ), или тиреотропин, активирует функцию щитовидной железы, вызывает гиперплазию ее железистой ткани, стимулирует выработку тироксина и трийодтиронина. Образование тиреотропина стимулируется тиреолиберином гипоталамуса, а угнетается соматостатином. Секреция тиреолиберина и тиреотропина регулируется йодсодержащими гормонами щитовидной железы по механизму обратной связи. Секреция тиреотропина усиливается также при охлаждении организма, что приводит к повышению выработки гормонов щитовидной железы и повышению тепла. Глюкокортикоиды тормозят продукцию тиреотропина. Секреция тиреотропина угнетается также при травме, боли, наркозе. Избыток тиреотропина проявляется гиперфункцией щитовидной железы, клинической картиной тиреотоксикоза. Фолликулостимулирующий гормон (ФСГ) и лютеинизирующий гормон (ЛГ)Фолликулостимулирующий гормон (ФСГ), или фоллитропин, вызывает рост и созревание фолликулов яичников и их подготовку к овуляции. У мужчин под влиянием ФСГ происходит образование сперматозоидов. Лютеинизирующий гормон (ЛГ), или лютропин, способствует разрыву оболочки созревшего фолликула, т.е. овуляции и образованию желтого тела. ЛГ стимулирует образование женских половых гормонов — эстрогенов. У мужчин этот гормон способствует образованию мужских половых гормонов — андрогенов. Секреция ФСГ и ЛС регулируется гонадолиберином гипоталамуса. Образование гонадолиберина, ФСГ и ЛГ зависит от уровня эстрогенов и андрогенов и регулируется по механизму обратной связи. Гормон аденогипофиза пролактин угнетает продукцию гонадотропных гормонов. Тормозное действие на выделение ЛГ оказывают глюкокортикоиды. Соматотропный гормон (СТГ)Соматотропный гормон (СТГ), или соматотропин, или гормон роста, принимает участие в регуляции процессов роста и физического развития. Стимуляция процессов роста обусловлена способностью соматотропина усиливать образование белка в организме, повышать синтез РНК, усиливать транспорт аминокислот из крови в клетки. Наиболее ярко влияние гормона выражено на костную и хрящевую ткани. Действие соматотропина происходит посредством «соматомединов», которые образуются в печени под влиянием соматотропина. Соматотропин влияет на углеводный обмен, оказывая инсулиноподобное действие. Гормон усиливает мобилизацию жира из депо и использование его в энергетическом обмене. Продукция соматотропина регулируется соматолиберином и соматостатином гипоталамуса. Снижение содержания глюкозы и жирных кислот, избыток аминокислот в плазме крови также приводят к увеличению секреции соматотропина. Вазопрессин, эндорфин стимулируют продукцию соматотропина. Если гиперфункция передней доли гипофиза проявляется в детском возрасте, то это приводит к усиленному пропорциональному росту в длину — гигантизму. Если гиперфункция возникает у взрослого человека, когда рост тела в целом уже завершен, наблюдается увеличение лишь тех частей тела, которые еще способны расти. Это пальцы рук и ног, кисти и стопы, нос и нижняя челюсть, язык, органы грудной и брюшной полостей. Это заболевание называется акромегалией. Причиной являются доброкачественные опухоли гипофиза. Гипофункция передней доли гипофиза в детстве выражается в задержке роста — карликовости («гипофизарный нанизм»). Умственное развитие не нарушено. Соматотропин обладает видовой специфичностью. ПролактинПролактин стимулирует рост молочных желез и способствует образованию молока. Гормон стимулирует синтез белка — лактальбумина, жиров и углеводов молока. Пролактин стимулирует также образование желтого тела и выработку им прогестерона. Влияет на водно-солевой обмен организма, задерживая воду и натрий в организме, усиливает эффекты альдостерона и вазопрессина, повышает образование жира из углеводов. Образование пролактина регулируется пролактолиберином и пролактостатином гипоталамуса. Установлено также, что стимуляцию секреции пролактина вызывают и другие пептиды, выделяющиеся гипоталамусом: тиреолиберин, вазоактивный интестинальный полипептид (ВИП), ангиотензин II, вероятно, эндогенный опиоидный пептид В-эндорфин. Секреция пролактина усиливается после родов и рефлекторно стимулируется при кормлении грудью. Эстрогены стимулируют синтез и секрецию пролактина. Угнетает продукцию пролактина дофамин гипоталамуса, который, вероятно, также тормозит клетки гипоталамуса, секретирующие гонадолиберин, что приводит к нарушению менструального цикла — лактогенной аменорее. Избыток пролактина наблюдается при доброкачественной аденоме гипофиза (гиперпролактинемическая аменорея), при менингитах, энцефалитах, травмах мозга, избытке эстрогенов, при применении некоторых противозачаточных средств. К его проявлениям относятся выделение молока у некормящих женщин (галакторея) и аменорея. Лекарственные вещества, блокирующие рецепторы дофамина (особенно часто психотропного действия),также приводят к повышению секреции пролактина, следствием чего могут быть галакторея и аменорея. Гормоны задней доли гипофизаЭти гормоны образуются в гипоталамусе. В нейрогипофизе происходит их накопление. В клетках супраоптического и паравентрикулярного ядер гипоталамуса осуществляется синтез окситоцина и антидиуретического гормона. Синтезированные гормоны путем аксонального транспорта с помощью белка — переносчика нейрофизина по гипоталамо-гипофизарному тракту — транспортируются в заднюю долю гипофиза. Здесь происходит депонирование гормонов и в дальнейшем выделение в кровь. Антидиуретический гормон (АДГ)Антидиуретический гормон (АДГ), или вазопрессин, осуществляет в организме 2 основные функции. Первая функция заключается в его антидиуретическом действии, которое выражается в стимуляции реабсорбции воды в дистальном отделе нефрона. Это действие осуществляется благодаря взаимодействию гормона с вазопрессиновыми рецепторами типа V-2, что приводит к повышению проницаемости стенки канальцев и собирательных трубочек для воды, ее реабсорбции и концентрированию мочи. В клетках канальцев происходит также активация гиалуронидазы, что приводит к усилению деполимеризации гиалуроновой кислоты, в результате чего повышается реабсорбция воды и увеличивается объем циркулирующей жидкости. В больших дозах (фармакологических) АДГ суживает артериолы, в результате чего повышается артериальное давление. Поэтому его также называют вазопрессином. В обычных условиях при его физиологических концентрациях в крови это действие не имеет существенного значения. Однако при кровопотере, болевом шоке происходит увеличение выброса АДГ. Сужение сосудов в этих случаях может иметь адаптивное значение. Образование АДГ усиливается при повышении осмотического давления крови, уменьшении объема внеклеточной и внутриклеточной жидкости, снижении артериального давления, при активации ренин-ангиотензиновой системы и симпатической нервной системы. При недостаточности образования АДГ развивается несахарный диабет, или несахарное мочеизнурение, который проявляется выделением больших количеств мочи (до 25 л в сутки) низкой плотности, повышенной жаждой. Причинами несахарного диабета могут быть острые и хронические инфекции, при которых поражается гипоталамус (грипп, корь, малярия), черепно-мозговые травмы, опухоль гипоталамуса. Избыточная секреция АДГ ведет, напротив, к задержке воды в организме. ОкситоцинОкситоцин избирательно действует на гладкую мускулатуру матки, вызывая ее сокращения при родах. На поверхностной мембране клеток существуют специальные окситоциновые рецепторы. Во время беременности окситоцин не повышает сократительную активность матки, но перед родами под влиянием высоких концентраций эстрогенов резко возрастает чувствительность матки к окситоцину. Окситоцин участвует в процессе лактации. Усиливая сокращения миоэпителиальных клеток в молочных железах, он способствует выделению молока. Увеличение секреции окситоцина происходит под влиянием импульсов от рецепторов шейки матки, а также механорецепторов сосков грудной железы при кормлении грудью. Эстрогены усиливают секрецию окситоцина. Функции окситоцина в мужском организме изучены не достаточно. Считают, что он является антагонистом АДГ. Недостаток продукции окситоцина вызывает слабость родовой деятельности. Гормоны гипофизаВ гипофизе синтезируется ряд биологически активных гормонов белковой и пептидной природы, оказывающих стимулирующий эффект на различные физиологические и биохимические процессы в тканях-мишенях (табл. 1). В зависимости от места синтеза различают гормоны передней, задней и промежуточной долей гипофиза. В передней доле вырабатываются в основном белковые и полипептидные гормоны, называемые тропными гормонами, или тропинами, вследствие их стимулирующего действия на ряд других эндокринных желез. В частности, гормон, стимулирующий секрецию гормонов щитовидной железы, получил название «тиротропин».
В последние годы из ткани мозга животных было выделено более 50 пептидов, получивших название нейропептидов и определяющих поведенческие реакции. Показано, что эти вещества влияют на некоторые формы поведения, процессы обучения и запоминания, регулируют сон и снимают, подобно морфину, боль. Так, выделенный β-эндорфин (31 аминокислотный остаток с выясненной последовательностью) оказался почти в 30 раз активнее морфина в качестве обезболивающего средства. Ряд других пептидов оказывает снотворное действие, а 16-членный пептид, вызывающий у крыс страх темноты, был назван скотофобином. Выделен полипептид амелетин, который, наоборот, отучает крыс бояться резкого звука электрического звонка. Работы в этом направлении интенсивно ведутся во многих лабораториях. Вполне возможно, что скоро будут выделены и соответственно синтезированы искусственно для каждой формы поведения соответствующие нейропептиды, включая пептиды памяти. Далее приводятся данные о структуре и функциях важнейших гормонов гипофиза и других желез внутренней секреции, имеющих белковую и пептидную природу. Вазопрессин и окситоцинГормоны вазопрессин и окситоцин синтезируются рибосомальным путем, причем одновременно в гипоталамусе синтезируются 3 белка: нейрофизин I, II и III, функция которых заключается в нековалентном связывании окситоцина и вазопрессина и транспорте этих гормонов в нейросекреторные гранулы гипоталамуса. Далее в виде комплексов нейрофизин–гормон они мигрируют вдоль аксона и достигают задней доли гипофиза, где откладываются про запас; после диссоциации комплекса свободный гормон секретируется в кровь. Нейрофизины также выделены в чистом виде, и выяснена первичная структура двух из них (92 из 97 аминокислотных остатков соответственно); это богатые цистеином белки, содержащие по семь дисульфидных связей. Химическое строение обоих гормонов было расшифровано классическими работами В. дю Виньо и сотр., впервые выделивших эти гормоны из задней доли гипофиза и осуществивших их химический синтез. Оба гормона представляют собой нонапептиды следующего строения: 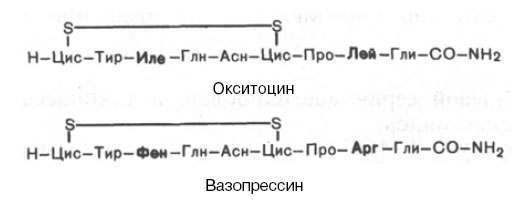 Вазопрессин отличается от окситоцина двумя аминокислотами: он содержит в положении 3 от N-конца фенилаланин вместо изолейцина и в положении 8 – аргинин вместо лейцина. Указанная последовательность 9 аминокислот характерна для вазопрессина человека, обезьяны, лошади, крупного рогатого скота, овцы и собаки. В молекуле вазопрессина из гипофиза свиньи вместо аргинина в положении 8 содержится лизин, отсюда название «лизин-вазопрессин». У всех позвоночных, за исключением млекопитающих, идентифицирован, кроме того, вазотоцин. Этот гормон, состоящий из кольца с S—S мостиком окситоцина и боковой цепью вазопрессина, был синтезирован химически В. дю Виньо задолго до выделения природного гормона. Высказано предположение, что эволюционно все нейрогипофизарные гормоны произошли от одного общего предшественника, а именно аргинин-вазотоцина, из которого путем одиночных мутаций триплетов генов образовались модифицированные гормоны. Основной биологический эффект окситоцина у млекопитающих связан со стимуляцией сокращения гладких мышц матки при родах и мышечных волокон вокруг альвеол молочных желез, что вызывает секрецию молока. Вазопрессин стимулирует сокращение гладких мышечных волокон сосудов, оказывая сильное вазопрессорное действие, однако основная роль его в организме сводится к регуляции водного обмена, откуда его второе название антидиуретического гормона. В небольших концентрациях (0,2 нг на 1 кг массы тела) вазопрессин оказывает мощное антидиуретическое действие – стимулирует обратный ток воды через мембраны почечных канальцев. В норме он контролирует осмотическое давление плазмы крови и водный баланс организма человека. При патологии, в частности атрофии задней доли гипофиза, развивается несахарный диабет – заболевание, характеризующееся выделением чрезвычайно больших количеств жидкости с мочой. При этом нарушен обратный процесс всасывания воды в канальцах почек. Относительно механизма действия нейрогипофизарных гормонов известно, что гормональные эффекты, в частности вазопрессина, реализуются через аденилатциклазную систему. Однако конкретный механизм действия вазопрессина на транспорт воды в почках пока остается неясным. Меланоцитстимулирующие гормоны (МСГ, меланотропины)Меланотропины синтезируются и секретируются в кровь промежуточной долей гипофиза. Выделены и расшифрованы первичные структуры двух типов гормонов – α- и β-меланоцитстимулирующие гормоны (α-МСГ и β- МСГ). Оказалось, чо у всех обследованных животных α-МСГ состоит из 13 остатков аминокислот, расположенных в одинаковой последовательности: СН3-СО-NH-Сер–Тир–Сер–Мет–Глу–Гис–Фен–Арг–Трп–Гли–Лиз–Про–Вал-СО-NН2 В α-МСГ N-концевой серин ацетилирован, а С-концевая аминокислота представлена валинамидом. Состав и структура β-МСГ оказались более сложными. У большинства животных молекула β-МСГ состоит из 18 остатков аминокислот; кроме того, имеются видовые различия, касающиеся природы аминокислоты в положениях 2, 6 и 16 полипептидной цепи гормона. β-МСГ, выделенный из промежуточной доли гипофиза человека, оказался 22-членным пептидом, удлиненным на 4 аминокислотных остатка с N-конца: Н-Ала–Глу–Лиз–Лиз–Асп–Глу–Гли–Про–Тир–Aрг–Мет–Глу–Гис–Фен– –Арг–Трп–Гли–Сер–Про–Про–Лиз–Асп-ОН Физиологическая роль меланотропинов заключается в стимулировании меланиногенеза у млекопитающих и увеличении количества пигментных клеток (меланоцитов) в кожных покровах земноводных. Возможно также влияние МСГ на окраску меха и секреторную функцию сальных желез у животных. Адренокортикотропный гормон (АКТГ, кортикотропин)Еще в 1926 г. было установлено, что гипофиз оказывает стимулирующее влияние на надпочечники, повышая секрецию гормонов коркового вещества. Накопленные к настоящему времени данные свидетельствуют, что этим свойством наделен АКТГ, вырабатываемый базофильными клетками аденогипофиза. АКТГ, помимо основного действия – стимуляции синтеза и секреции гормонов коры надпочечников, обладает жиромобилизующей и меланоцитстимулирующей активностью. Молекула АКТГ у всех видов животных содержит 39 аминокислотных остатков. Первичная структура АКТГ свиньи и овцы была расшифрована еще в 1954–1955 гг. Приводим уточненное строение АКТГ человека: Н-Сер–Тир–Сер–Мет–Глу–Гис–Фен–Арг–Трп–Гли–Лиз–Про–Вал–Гли–Лиз–Лиз–Aрг–Aрг–Про–Вал–Лиз–Вал–Тир–Про–Асп–Ала–Гли–Глу–Асп–Глн–Сер–Ала–Глу–Ала–Фен–Про–Лей–Глу–Фен-ОН Различия в структуре АКТГ овцы, свиньи и быка касаются только природы 31-го и 33-го остатков аминокислот, однако все они наделены почти одинаковой биологической активностью, как и АКТГ гипофиза человека. В молекуле АКТГ, как и других белковых гормонов, хотя и не открыты активные центры наподобие активных центров ферментов, однако предполагается наличие двух активных участков пептидной цепи, один из которых ответствен за связывание с соответствующим рецептором, другой – за гормональный эффект. Данные о механизме действия АКТГ на синтез стероидных гормонов свидетельствуют о существенной роли аденилатциклазной системы. Предполагают, что АКТГ вступает во взаимодействие со специфическими рецепторами на внешней поверхности клеточной мембраны (рецепторы представлены белками в комплексе с другими молекулами, в частности с сиаловой кислотой). Сигнал затем передается на фермент аденилатциклазу, расположенную на внутренней поверхности клеточной мембраны, которая катализирует распад АТФ и образование цАМФ. Последний активирует протеинкиназу, которая в свою очередь с участием АТФ осуществляет фосфорилирование холинэстеразы, превращающей эфиры холестерина в свободный холестерин, который поступает в митохондрии надпочечников, где содержатся все ферменты, катализирующие превращение холестерина в кортикостероиды. Соматотропный гормон (СТГ, гормон роста, соматотропин)Гормон роста был открыт в экстрактах передней доли гипофиза еще в 1921 г., однако в химически чистом виде получен только в 1956–1957 гг. СТГ синтезируется в ацидофильных клетках передней доли гипофиза; концентрация его в гипофизе составляет 5–15 мг на 1 г ткани, что в 1000 раз превышает концентрацию других гормонов гипофиза. К настоящему времени полностью выяснена первичная структура белковой молекулы СТГ человека, быка и овцы. СТГ человека состоит из 191 аминокислоты и содержит две дисульфидные связи; N- и С-концевые аминокислоты представлены фенилаланином. СТГ обладает широким спектром биологического действия. Он влияет на все клетки организма, определяя интенсивность обмена углеводов, белков, липидов и минеральных веществ. Он усиливает биосинтез белка, ДНК, РНК и гликогена и в то же время способствует мобилизации жиров из депо и распаду высших жирных кислот и глюкозы в тканях. Помимо активации процессов ассимиляции, сопровождающихся увеличением размеров тела, ростом скелета, СТГ координирует и регулирует скорость протекания обменных процессов. Кроме того, СТГ человека и приматов (но не других животных) обладает измеримой лактогенной активностью. Предполагают, что многие биологические эффекты этого гормона осуществляются через особый белковый фактор, образующийся в печени под влиянием гормона. Этот фактор был назван сульфирующим или тимидиловым, поскольку он стимулирует включение сульфата в хрящи, тимидина – в ДНК, уридина – в РНК и пролина – в коллаген. По своей природе этот фактор оказался пептидом с мол. массой 8000. Учитывая его биологическую роль, ему дали наименование «соматомедин», т.е. медиатор действия СТГ в организме. СТГ регулирует процессы роста и развития всего организма, что подтверждается клиническими наблюдениями. Так, при гипофизарной карликовости (патология, известная в литературе как пангипопитуитаризм; связана с врожденным недоразвитием гипофиза) отмечается пропорциональное недоразвитие всего тела, в том числе скелета, хотя существенных отклонений в развитии психической деятельности не наблюдается. У взрослого человека также развивается ряд нарушений, связанных с гипо- или гиперфункцией гипофиза. Известно заболевание акромегалия (от греч. akros – конечность, megas – большой), характеризующееся непропорцио- нально интенсивным ростом отдельных частей тела, например рук, ног, подбородка, надбровных дуг, носа, языка, и разрастанием внутренних органов. Болезнь вызвана, по-видимому, опухолевым поражением передней доли гипофиза. Лактотропный гормон (пролактин, лютеотропный гормон)Пролактин считается одним из наиболее «древних» гормонов гипофиза, поскольку его удается обнаружить в гипофизе низших наземных животных, у которых отсутствуют молочные железы, а также получить лактогенный эффект у млекопитающих. Помимо основного действия (стимуляция развития молочных желез и лактации), пролактин имеет важное биологическое значение – стимулирует рост внутренних органов, секрецию желтого тела (отсюда его второе название «лютеотропный гормон»), оказывает ренотропное, эритропоэтическое и гипергликемическое действие и др. Избыток пролактина, образующийся обычно при наличии опухолей из секретирующих пролактин клеток, приводит к прекращению менструаций (аменорея) и увеличению молочных желез у женщин и к импотенции – у мужчин. Расшифрована структура пролактина из гипофиза овцы, быка и человека. Это крупный белок, представленный одной полипептидной цепью с тремя дисульфидными связями, состоящий из 199 аминокислотных остатков. Видовые отличия в последовательности аминокислот касаются по существу 2–3 аминокислотных остатков. Раньше оспаривалось мнение о секреции лактотропина в гипофизе человека, поскольку предполагали, что его функцию якобы выполняет соматотропин. В настоящее время получены убедительные доказательства существования пролактина человека, хотя в гипофизе его содержится значительно меньше, чем гормона роста. В крови женщин уровень пролактина резко повышается перед родами: до 0,2 нг/л против 0,01 нг/л в норме. Тиреотропный гормон (ТТГ, тиротропин)В отличие от рассмотренных пептидных гормонов гипофиза, представленных в основном одной полипептидной цепью, тиротропин является сложным гликопротеином и содержит, кроме того, по две α- и β-субъединицы, которые в отдельности биологической активностью не обладают: мол. масса его около 30000. Тиротропин контролирует развитие и функцию щитовидной железы и регулирует биосинтез и секрецию в кровь тиреоидных гормонов. Полностью расшифрована первичная структура α- и β-субъединиц тиротропина быка, овцы и человека: &lpha;-субъединица, содержащая 96 аминокислотных остатков, имеет одинаковую аминокислотную последовательность во всех изученных ТТГ и во всех лютеинизирующих гормонах гипофиза; β-субъединица тиротропина человека, содержащая 112 аминокислотных остатков, отличается от аналогичного полипептида в ТТГ крупного рогатого скота аминокислотными остатками и отсутствием С-концевого метионина. По-этому многие авторы специфические биологические и иммунологические свойства гормона объясняют наличием β-субъединицы ТТГ в комплексе с α-субъединицей. Предполагают, что действие тиротропина осуществляется, подобно действию других гормонов белковой природы, посредством связывания со специфическими рецепторами плазматических мембран и активирования аденилатциклазной системы. Гонадотропные гормоны (гонадотррпины)К гонадотропинам относятся фолликулостимулирующий гормон (ФСГ, фоллитропин) и лютеинизирующий гормон (ЛГ, лютропин), или гормон, стимулирующий интерстициальные клетки. Оба гормона синтезируются в передней доле гипофиза и являются, как и тиротропин, сложными белками – гликопротеинами с мол. массой 25000. Они регулируют стероидо- и гаметогенез в половых железах. Фоллитропин вызывает созревание фолликулов в яичниках у самок и сперматогенез – у самцов. Лютропин у самок стимулирует секрецию эстрогенов и прогестерона, как и разрыв фолликулов с образованием желтого тела, а у самцов – секрецию тестостерона и развитие интерстициальной ткани. Биосинтез гонадотропинов, как было отмечено, регулируется гипоталамическим гормоном гонадолиберином. Химическая структура молекулы лютропина расшифрована полностью. Лютропин состоит из двух α- и β-субъединиц. Структура α-субъединиц гормона у большинства животных совпадает. Так, у овцы она содержит 96 аминокислотных остатков и 2 углеводных радикала. У человека α-субъединица гормона укорочена на 7 аминокислотных остатков с N-конца и отличается природой 22 аминокислот. Расшифрована также последовательность аминокислот в β-субъединицах лютропина свиньи и человека. α- и β-Субъединицы в отдельности лишены биологической активности (по аналогии с большинством субъединиц ферментов). Только их комплекс, образование которого, вероятнее всего, предопределено первичной структурой их, приводит к формированию биологически активной макромолекулярной структуры за счет гидрофобных взаимодействий. Липотропные гормоны (ЛТГ, липотропины)Среди гормонов передней доли гипофиза, структура и функция которых выяснены в последнее десятилетие, следует отметить липотропины, в частности β- и γ-ЛТГ. Наиболее подробно изучена первичная структура β-липотропина овцы и свиньи, молекулы которого состоят из 91 аминокислотного остатка и имеют существенные видовые различия в последовательности аминокислот. К биологическим свойствам β-липотропина относятся жиромобилизующее действие, кортикотропная, меланоцитстимулирующая и гипокальциемическая активность и, кроме того, инсулиноподобный эффект, выражающийся в повышении скорости утилизации глюкозы в тканях. Предполагают, что липотропный эффект осуществляется через систему аденилатциклаза–цАМФ–протеинкиназа, завершающей стадией действия которой является фосфорилирование неактивной триацилглицерол-липазы. Этот фермент после активирования расщепляет нейтральные жиры на диацилглицерол и высшую жирную кислоту. Перечисленные биологические свойства обусловлены не β-липотропином, оказавшимся лишенным гормональной активности, а продуктами его распада, образующимися при ограниченном протеолизе. Оказалось, что в ткани мозга и в промежуточной доле гипофиза синтезируются биологически активные пептиды, наделенные опиатоподобным действием. Приводим структуры некоторых из них: Н–Тир–Гли–Гли–Фен–Мет–ОН
Н–Тир–Гли–Гли–Фен–Лей–ОН
Н–Тир–Гли–Гли–Фен–Мет–Тре–Сер–Глу–Лиз–Сер–Глн–Тре–Про–Лей–Вал–Тре–Лей–Фен–Лиз–Асн–Ала–Иле–Вал–Лиз–Асн–Ала–Гис–Лиз–Лиз–Гли–Глн–ОН
Общим типом структуры для всех трех соединений является тетрапептидная последовательность на N-конце. Доказано, что β-эндорфин (31 АМК) образуется путем протеолиза из более крупного гипофизарного гормона β-липотропина (91 АМК); последний вместе с АКТГ образуется из общего предшественника – прогормона, названного проопиокортином (является, таким образом, препрогормоном), имеющим молекулярную массу 29 кДа и насчитывающим 134 аминокислотных остатка. Биосинтез и освобождение проопиокортина в передней доле гипофиза регулируется кортиколиберином гипоталамуса. В свою очередь из АКТГ и β-липотропина путем дальнейшего процессинга, в частности ограниченного протеолиза, образуются соответственно α- и β-меланоцитстимулирующие гормоны (α- и β-МСГ). С помощью техники клонирования ДНК, а также метода определения первичной структуры нуклеиновых кислот Сенджера в ряде лабораторий была раскрыта нуклеотидная последовательность мРНК–предшественника проопиокортина. Эти исследования могут служить основой для целенаправленного получения новых биологически активных гормональных лечебных препаратов. Ниже представлены пептидные гормоны, образующиеся из β-липотропина путем специфического протеолиза.
Учитывая исключительную роль β-липотропина как предшественника перечисленных гормонов, приводим первичную структуру β-липотропина свиньи (91 аминокислотный остаток): Н–Глу–Лей–Ала–Гли–Ала–Про–Про–Глу–Про–Ала–Aрг–Асп–Про–Глу–Ала–Про–Ала–Глу–Гли–Ала–Ала–Ала–Aрг–Ала–Глу–Лей–Глу–Тир–Гли–Лей–Вал–Ала–Глу–Ала–Глу–Ала–Ала–Глу–Лиз–Лиз–Асп–Глу–Гли–Про–Тир–Лиз–Мет–Глу–Гис–Фен–Арг–Трп–Гли–Сер–Про–Про–Лиз–Асп–Лиз–Aрг–Тир–Гли–Гли–Фен–Мет–Тре–Сер–Глу–Лиз–Сер–Глн–Тре–Про–Лей–Вал–Тре–Лей–Фен–Лиз–Асн–Ала–Иле–Вал–Лиз–Асн–Ала–Гис–Лиз–Лиз–Гли–Глн–ОН Повышенный интерес к указанным пептидам, в частности энкефалинам и эндорфинам, диктуется их необычайной способностью, подобно морфину, снимать болевые ощущения. Эта область исследования – поиск новых природных пептидных гормонов и(или) их направленный биосинтез – является интересной и многообещающей для развития физиологии, нейробиологии, неврологии и клиники. |
|
||||||||||||||||||||||||||||||||||||||