ALLHORMON.NAROD.RU
Все о гормона - сайт посвященные эндокринной системе человека.
|
Главная |
Добавить в избранное
ГипоталамусГипоталамус служит местом непосредственного взаимодействия высших отделов ЦНС и эндокринной системы. Природа связей, существующих между ЦНС и эндокринной системой, стала проясняться в последние десятилетия, когда из гипоталамуса были выделены первые гуморальные факторы, оказавшиеся гормональными веществами с чрезвычайно высокой биологической активностью. Потребовалось немало труда и экспериментального мастерства, чтобы доказать, что эти вещества образуются в нервных клетках гипоталамуса, откуда по системе портальных капилляров достигают гипофиза и регулируют секрецию гипофизарных гормонов, точнее их освобождение. Эти вещества получили сначала наименование нейрогормонов, а затем рилизинг-факторов (от англ.release – освобождать), или либеринов. Вещества с противоположным действием, т.е. угнетающие освобождение гипофизарных гормонов, стали называть ингибирующими факторами, или статинами. Таким образом, гормонам гипоталамуса принадлежит ключевая роль в физиологической системе гормональной регуляции многосторонних биологических функций отдельных органов, тканей и целостного организма. Анатомическое строениеГипоталамус - главный подкорковый центр регуляции вегетативно-висцеральных и эндокринных функций. Он занимает вентральную часть промежуточного мозга и располагается ниже подбугорной борозды, sulcus hypothalamicus. Делится на зрительную и обонятельную части, pars optica et pars olfactoria. В настоящее время в гипоталамусе описано 32-48 ядер, являющимися высшими вегетативными центрами, регулирующими все виды обмена веществ, терморегуляцию и т. д.
Некоторые ядра обладают нейросекреторной функцией (супраоптические, паравентрикулярные, дугообразные и вентромедиальные). Гистологическое строениеГипоталамус - участок промежуточного мозга, содержащий особые нейросекреторные ядра, клетки которых вырабатывают и секретируют в кровь нейрогормоны. Эти клетки получают афферентные импульсы из других частей нервной системы, а их аксоны оканчиваются на кровеносных сосудах (аксо-вазальные синапсы). Нейросекреторные клетки - отростчатой формы, с крупным везикулярным ядром, хорошо заметным ядрышком и базофильной цитоплазмой, содержащей развитую грЭПС и крупный комплекс Гольджи, от которого отделяются нейросекреторные гранулы (рис. 1). Гранулы транспортируются со скоростью около 1-4 мм/ч по аксону вдоль центрального пучка микротрубочек и микрофиламентов, а местами накапливаются в больших количествах, растягивая аксон. Самые крупные из таких участков хорошо видны под световым микроскопом и называются накопительными нейросекреторными тельцами (Херринга); в них сосредоточено до 60% всего нейросекрета, лишь около 30% находится в области терминалей. Терминали (аксо-вазальные синапсы) характеризуются присутствием, помимо гранул, многочисленных светлых пузырьков (осуществляют возврат мембраны после экзоцитоза). 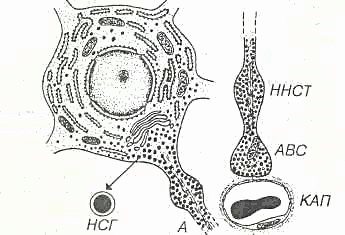 Рис. 1. Ультраструктурная организация нейросекреторных клеток.А - аксон, ABC - аксо-вазапьный синапс, КАП - капилляр, ННСТ - накопительное нейросекреторное тельце, НСГ - нейросекреторные гранулы. Нейросекреторные ядра гипоталамуса в зависимости от размеров клеток и их функциональных особенностей разделяют на крупно- и мелкоклеточные. 1. Крупноклеточные ядра образованы клеточными телами, которые в 2-3 раза крупнее, чем в других отделах гипоталамуса; к ним относятся супраоптическое (СОЯ) и паравентрикулярное (ПВЯ) ядра. СОЯ в 3-4 раза объемнее ПВЯ; ПВЯ лишь в центральных участках построено по типу крупноклеточного, а в периферических отделах образовано мелкими нейросекреторными клетками. Аксоны клеток СОЯ и ПВЯ покидают гипоталамус и, в составе гипоталамо-гипофизарного тракта пересекая гемато-энцефалическип барьер, проникают в заднюю долю гипофиза, где образуют терминали на капиллярах (рис. 2). Крупноклеточные ядра секретируют антидиуретический гормон (АДГ) или вазопрессин (ВП), и окситоцин. Эти гормоны вырабатываются разными клетками. У человека АДГ образуется, главным образом, в СОЯ, а окситоцин - в ПВЯ. АДГ и окситоцин синтезируются в грЭПС (в виде крупной молекулы прогормона) и переносятся в комплекс Гольджи, где упаковываются в гранулы. Процессинг продукта (завершается лишь в ходе транспорта гранул в аксоне) приводит к освобождению активного гормона и нейрофизина - белка с неясной функцией (ранее считали переносчиком нейрогормонов). 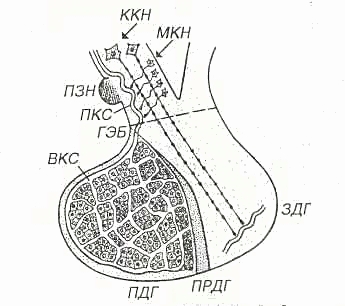 Рис. 2. Схема строения гипоталамо-гипофизарной нейросекреторной системы.ПДГ - передняя доля гипофиза, ПРДГ - промежуточная доля гипофиза, ЗДГ - задняя доля гипофиза, ККН - крупноклеточные нейроны, МКН - мелкоклеточные нейроны ПЗН - перекрест зрительных нервов, ГЭБ - гемато-энцефалический барьер, ПКС - первичная капиллярная сеть, ВКС - вторичная капиллярная сеть. 2. Мелкоклеточные ядра вырабатывают ряд гипофизотропных факторов, которые усиливают (рилизинг факторы или либерины) или угнетают (ингабирующие факторы или статины) выработку гормонов клетками передней доли, попадая к ним по воротной системе сосудов. Аксоны нейросекреторных клеток этих ядер образуют терминалы на первичной капиллярной сети в срединном возвышении (нейрогемальной контактной зоне). Эта сеть далее собирается в воротные вены, проникающие в переднюю долю гипофиза и распадающиеся на вторичную сеть капилляров между тяжами железистых клеток - аденоцитов (см. рис. 2). Физиология гипоталамусаОсновные связи между нервной и эндокринной системами регуляции осуществляются посредством взаимодействия гипоталамуса и гипофиза (рис. 3). Нервные импульсы, приходящие в гипоталамус, активируют секрецию так называемых рилизинг-факторов (либеринов и статинов): тиреолиберина, соматолиберина, пролактолиберина, гонадолиберина и кортиколиберина, а также соматостатина и пролактостатина. Мишенью для либеринов и статинов, секретируемых гипоталамусом, является гипофиз. Каждый из либеринов взаимодействует с определенной популяцией клеток гипофиза и вызывает в них синтез соответствующих тропинов: тиреотропина, соматотропного гормона (соматотропин — гормон роста), пролактина, гонадотропного гормона, (гонадотропины — лютеинизирующий и фолликулостимулирующий), а также адренокортикотропного гормона (АКТГ, кортикотропин). Статины оказывают на гипофиз влияние, противоположное действию либеринов, — подавляют секрецию тропинов. Тропины, секретируемые гипофизом, поступают в общий кровоток и, попадая на соответствующие железы, активируют в них секреторные процессы. 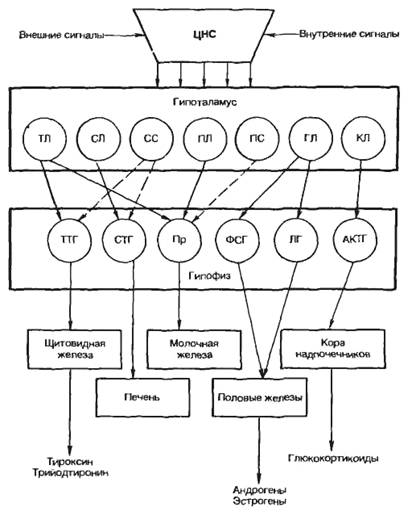 Рис 3. Регуляция активности эндокринных желез центральной нервной системой при участии гипоталамуса и гипофиза. ТЛ — тиреолиберин; СЛ — соматалиберин; СС — соматостатин; ПЛ — пролактолиберин; ГЛ — гонадолиберин; КЛ — кортиколиберин; ТТГ — тиреотропный гормон; СТГ — соматотропный гормон (гормон роста); Пр — пролактин; ФСГ — фолликулостимулирующий гормон; ЛГ — лютеиниэирующий гормон; АКТГ — адренокортикотропный гормон. Сплошными стрелками обозначено активирующее, пунктирными — ингибирующее влияние. Регуляция деятельности гипофиза и гипоталамуса, кроме сигналов, идущих «сверху вниз», осуществляется гормонами «исполнительных» желез (рис. 3). Эти «обратные» сигналы поступают в гипоталамус и затем передаются в гипофиз, что приводит к изменению секреции соответствующих тропинов. После удаления или атрофии эндокринной железы стимулируется секреция соответствующего тропного гормона; при гиперфункции железы секреция соответствующего тропина подавляется. 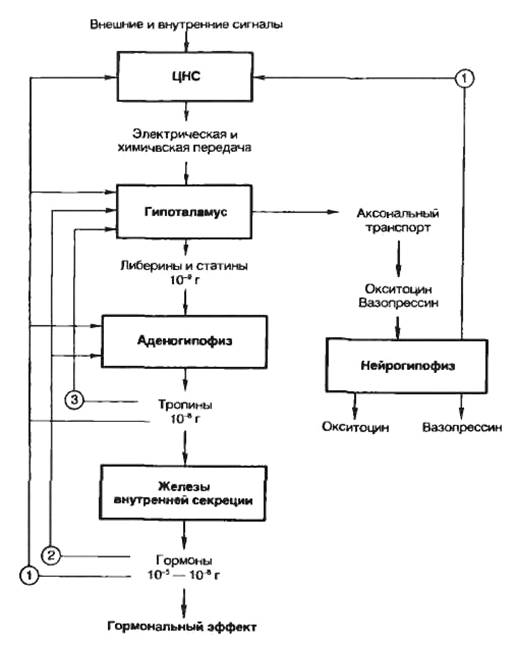 Рис.4 Прямые и обратные связи в нейроэндокринной системе регуляции.1 — медленно развивающееся и продолжительное ингибирование секреции гормонов и нейромедиаторов, а также изменение поведения и формирование памяти; 2 — быстро развивающееся, но продолжительное ингибирование; 3 — кратковременное ингибирование. Обратные связи не только позволяют регулировать концентрацию гормонов в крови, но и участвуют в дифференцировке гипоталамуса в онтогенезе. Образование половых гормонов в женском организме происходит циклически, что объясняется циклической секрецией гонадотропных гормонов. Синтез этих гормонов контролируется гипоталамусом, образующим рилизинг-фактор этих тропинов (гонадолиберин). Если самке пересадить гипофиз самца, то пересаженный гипофиз начинает функционировать циклично. Половая дифференцировка гипоталамуса происходит под действием андрогенов. Если самца лишить половых желез, продуцирующих андрогены, то гипоталамус будет дифференцироваться по женскому типу. В железах внутренней секреции иннервированы, как правило, только сосуды, а эндокринные клетки изменяют свою биосинтетическую и секреторную активность лишь под действием метаболитов, кофакторов и гормонов, причем не только гипофизарных. Так, ангиотензин II стимулирует синтез и секрецию альдостерона. Отметим также, что некоторые гормоны гипоталамуса и гипофиза могут образовываться не только в этих тканях. Например, соматостатин (гормон гипоталамуса, ингибирующий образование и секрецию гормона роста) обнаружен также в поджелудочной железе, где он подавляет секрецию инсулина и глюкагона. Большинство нервных и гуморальных путей регуляции сходится на уровне гипоталамуса и благодаря этому в организме образуется единая нейроэндокринная регуляторная система. К клеткам гипоталамуса подходят аксоны нейронов, расположенных в коре больших полушарий и подкорковых образованиях. Эти аксоны секретируют различные нейромедиаторы, оказывающие на секреторную активность гипоталамуса как активирующее, так и тормозное влияние. Поступающие из мозга нервные импульсы гипоталамус «превращает» в эндокринные стимулы, которые могут быть усилены или ослаблены в зависимости от гуморальных сигналов, поступающих в гипоталамус от желез и тканей, подчиненных ему. Тропины, образующиеся в гипофизе, не только регулируют деятельность подчиненных желез, но и выполняют самостоятельные эндокринные функции. Например, пролактин оказывает лактогенное действие, а также тормозит процессы дифференцировки клеток, повышает чувствительность половых желез к гонадотропинам, стимулирует родительский инстинкт. Кортикотропин является не только стимулятором стероцдогенеза, но и активатором липолиза в жировой ткани, а также важнейшим участником процесса превращения в мозге кратковременной памяти в долговременную. Гормон роста может стимулировать активность иммунной системы, обмен липидов, сахаров и т. д. В задней доле гипофиза (нейрогипофиз) депонируются антидиуретический гормон (вазопрессин) и окситоцин (см. рис. 2). Первый вызывает задержку воды в организме и повышает тонус сосудов, второй стимулирует сокращение матки при родах и секрецию молока. Оба гормона синтезируются в гипоталамусе, затем транспортируются по аксонам в заднюю долю гипофиза, где депонируются и потом секретируются в кровь. Характер процессов, протекающих в ЦНС, во многом определяется состоянием эндокринной регуляции. Так, андрогены и эстрогены формируют половой инстинкт, многие поведенческие реакции. Очевидно, что нейроны, точно так же как и другие клетки нашего организма, находятся под контролем гуморальной системы регуляции. Нервная система, эволюционно более поздняя, имеет как управляющие, так и подчиненные связи с эндокринной системой. Эти две регуляторные системы дополняют друг друга, образуют функционально единый механизм, что обеспечивает высокую эффективность нейрогуморальной регуляции, ставит ее во главе систем, согласующих все процессы жизнедеятельности в многоклеточном организме. Гормоны гипоталамусаК настоящему времени в гипоталамусе открыто 7 стимуляторов и 3 ингибитора секреции гормонов гипофиза, а именно: кортиколиберин, тиролиберин, люлиберин, фоллилиберин, соматолиберин,пролактолиберин, меланолиберин, соматостатин, пролактостатин и меланостатин. В чистом виде выделено 5 гормонов, для которых установлена первичная структура, подтвержденная химическим синтезом. Большие трудности при получении гормонов гипоталамуса в чистом виде объясняются чрезвычайно низким содержанием их в исходной ткани. По химическому строению все гормоны гипоталамуса являются низкомолекулярными пептидами, так называемыми олигопептидами необычного строения, хотя точный аминокислотный состав и первичная структура выяснены не для всех. Приводим полученные к настоящему времени данные о химической природе шести из известных 10 гормонов гипоталамуса. 1. Тиролиберин (Пиро-Глу–Гис–Про–NH2):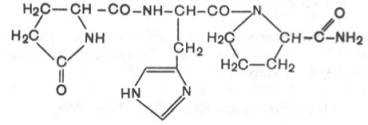 Тиролиберин представлен трипептидом, состоящим из пироглутаминовой (циклической) кислоты, гистидина и пролинамида, соединенных пептидными связями. В отличие от классических пептидов он не содержит свободных NH2- и СООН-групп у N- и С-концевых аминокислот. Обеспечивает высвобождение тиротропного гормона (ТТГ) из передней доли гипофиза. 2. Гонадолиберин является декапептидом, состоящим из 10 аминокислот последовательности: Обеспечивает высвобождение ФСГ, ЛГ и пролактина 3. Соматостатин является циклическим тетрадекапептидом (состоит из 14 аминокислотных остатков) :  Отличается этот гормон от двух предыдущих, помимо циклической структуры, тем, что не содержит на N-конце пироглутаминовой кислоты: дисульфидная связь образуется между двумя остатками цистеина в 3-м и 14-м положениях. Следует отметить, что синтетический линейный аналог соматостатина также наделен аналогичной биологической активностью, что свидетельствует о несущественности дисульфидного мостика природного гормона. Помимо гипоталамуса, соматостатин продуцируется нейронами центральной и периферической нервных систем, а также синтезируется в S-клетках панкреатических островков (островков Лангерганса) в поджелудочной железе и клетках кишечника. Он оказывает широкий спектр биологического действия; в частности, показано ингибирующее действие на синтез гормона роста в аденогипофизе, а также прямое тормозящее действие его на биосинтез инсулина и глюкагона в β- и α-клетках островков Лангерганса. 4. Соматолиберин. Он представлен 44 аминокислотными остатками с полностью раскрытой последовательностью. Биологической активностью соматолиберина наделен, кроме того, химически синтезированный декапептид: Н-Вал–Гис–Лей–Сер–Ала–Глу–Глн–Лиз–Глу–Ала-ОН. Этот декапептид стимулирует синтез и секрецию гормона роста гипофиза соматотропина. 5. Меланолиберин, химическая структура которого аналогична структуре открытого кольца гормона окситоцина (без трипептидной боковой цепи), имеет следующее строение: Н-Цис–Тир–Иле–Глн–Асн–Цис-ОН. 6. Меланостатин (меланотропинингибирующий фактор) представленили трипептидом: Пиро-Глу–Лей–Гли-NН 2 , или пентапептидом со следующей последовательностью: Пиро-Глу–Гис–Фен–Aрг–Гли–NН2. Необходимо отметить, что меланолиберин оказывает стимулирующее действие, а меланостатин, напротив, ингибирующее действие на синтез и секрецию меланотропина в передней доле гипофиза. |
|